




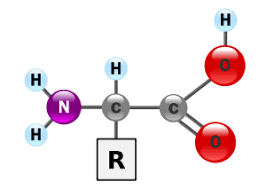
مقاله کامل یون
مقاله کامل یون
تنظيم تركيبات يوني و PH ميان سلولي :
توزيع و انتشار ويژه اي از يونها در سراسر غشاء پلاسماي سلولهاي يوكاريوتيك ewkaryotic وجود دارد كه از طريق مكانيزمهاي انتقال فعال يوني و نفوذپذيري يون و غشاء انتخابي انجام مي شود . اين حالت منجر به تفاوت پتاسين در سراسر غشاء در حدود 70mv در طي شرايط ساكن مي شود كه نزديك به پتانسيل ثابت براي پتاسيم است بنابراين گراديان براي سديم و كلسيم به داخل سلل هدايت مي شوند در حاليكه گراديان پتاسيم به سمت خارج است . منيزيم يونيزه شده تقريبا بطور مساوي و يكسان در سراسر غشاء پلاسمايي منتشر مي شود گرچه به علت پتانسيل منفي ساكن مقدار آن از ميزان ثابت الكتروشيميايي بيشتر است . محدوده فيزيولوژيكي اين يونها در بدنه خارج سلولي و داخل سلولي در جداول 6-1 ارائه شده است .
تعادل يون بهينه شده براي متابوليسم ، رشد معمولي و عملكرد سلولها لازم و اساسي است مكانيزمهاي موثر غلظت يون را در ميان طيف هاي باريك خود تنظيم و كنترل مي كنند به اين دليل سلولهاي يوكاريوت شامل كانالها و ناقلهاي يوني مختلفي در غشاء هستند علاوه بر اين مكانيزمها ارتباط نزديكي با تنظيم و كنترل حجم و PH درون سلولي دارند.
پتاسيم :
پتاسيم يكي از كاتيونهاي بسيار فراوان بدن است ، بالغ بر 3500 تا 4000 ميلي مول در كل است . مقدار كلي مرتبط با جرم چربي آزاد بدن است و در حدود 50-70 است . از كل پتاسيم بدن 98% در خارج سلول واقع شده است در حاليكه باقي مانده در بدنه هاي مختلف خارج سلولي (فاصله بافت و غشاء 40mmol – مابع ميان سلولي
35mmol ، پلاسما 40mmol ) اين گراديان پتاسيم براي تنظيم حجم سلول به علاوه براي تحريك پذيري سلولهاي عصبي و ماهيچه اي بسيار اهميت دارد .
پروتئينهاي غشاء تنظيم كننده پتاسيم:
توزيع و انتشار نامتعادل پتاسيم غالبا نتيجه فرايندهاي انتقال فعال At pox سديم – پتاسيم است . علاوه بر اين توزيع و انتشار آن بستگي به تراوش پذيري و نفوذپذيري غشاء پلاسما براي پتاسيم دارد كه متاثر از مسيرها و كنالهائي مختلف انتقال پتاسيم است . از اينرو تعادل پتاسيم درون سلولي بطور عالي از طريق ساختارهاي مختلف حاصل پتاسيم نظير پمپ هاي يوني ،كانالهاي پتاسيم و ناقلهاي پتاسيم تنظيم و كنترل مي شود .
جدول 6-1
توزيع يونها در سراسر غشاء پلاسمايي گلبولهاي قرمز خون
مهمترين مورد در خصوص ورزش در پاراگرافهاي بعدي آمده است
– كانالهاي پتاسيم :
كانالهاي پتاسيم بزرگترين و متنوع ترين زير مجموعه اي از كانالهاي يوني است در يك بررسي و تجديد نظر ،در كانال اصلي پتاسيم عبارتند از كانالهاي پتاسيم حساس به ولتاژ و كانالهاي يكسوساز پتاسيم داخلي هر دو نوع كانالها شامل چهار زنجيره يا زير واحد پپتید هستند كه به صورت همگن يا ناهمگن با هم در ارتباط هستند تا مسير نفوذ تراوش را در سراسر غشاء تشكيل دهند . زنجيره هاي پپتید در كانالهاي پتاسيم يکسو ساز در داخل سلول شامل دو مارپیچ در يك طرف غشاء با بخشي از آمينواسيدهاي كوچك ميان آنها مي باشد . اين وضعيت لوپ p ناميده مي شود . زيرا آن داخل غشاء بدون اينكه
كاملا آنرا قطع كرده باشد قرار دارد چنين ساختاري ويژگي مشخصه تمام انواع كانالهاي پتاسيم است .
كانالهاي پتاسيم حساس به ولتاژ
در اين نوع كانال (kv) چهار زنجيره اضافي پپتید جلوتر از دو حلقه مارپيچ ميان غشائي قرار دارد كه شامل واحدهاي حساس به ولتاژ هستند كانالهاي حساس به ولتاژ تفاوت زياد در حساسيت پذيري ولتاژ و فعال سازي انرژي جنبشي را نشان مي دهد . حداقل 22ژن مختلف داراي كد و رمز براي اين نوع كانالهاي در پستانداران شناسايي شده است . در صورتيكه انواع زيادي توسط عمليات پيوند و دايمر كردن نامشابه توليد شده اند . يك نمونه كانال پتاسيم یکسو ساز تاخيري است كه تقريبا در تمام غشاء هاي تحريك پذير وجود دارد . اين كانال به محض اينكه غشاء قطبش زدايي شد پس از يك تاخير كوتاه باز مي شود . بدين وسيله پتانسيل بازگرداننده به سطح ساكن بر مي گردد. گروه ديگر از كانالهاي پتاسيم ورودي ولتاژ توسط كانالهاي پتاسيم فعال شده كلسيم نمايان مي شوند . اين نوع از كانالها توسط افزايش و يا قطبش زدايي كلسيم فعال شده فعال مي شوند كانالهاي پتاسيم فعال شده كلسيم حداقل سه زير مجموعه به علت خاصيت رسانايي آنها تقسيم مي شوند . بزرگ (200-300ps) توسط 20-60ps و كوچك 10-14ps احتمال بازشدن كانال توسط ميزان زيادي از غلظت كلسيم درون سلولي درصد را 50mmol تا0/5 تنظيم مي شود كه تنوع سايت ها و مكانهاي اتصال كلسيم هاي تنظيم كننده را نشان مي دهد و نقش اين كانالها تنها تحت شرايط افزايش غلظت كلسيم درون سلولي در طي فعاليت هاي مداوم عضلاني و يا در ضميرهاي تحليل رفته مشخص مي شود.
كانالهاي پتاسيم یکسو ساز داخلي
اجزاء بين كانالها داراي يك خاصيت مشترك هستند و همه در تنظيم توسط مدولاتورهاي مختلف داخل سلولي و يا پيك هاي داخل سلولي نظير كلئوتيدها ، فسفرليپيدها ، يكفازها ، پلي آميدها ، PH و پروتئين G سهيم هستند كانالهاي پتاسيم یکسوكننده داخلي به خوبي و درستي تحريك پذيري غشاء را تنظيم مي كنند . باز شدن كانال پتاسيم غشاء و را نزدیک پتانسيل متعادل پتاسيم به توازن مي رساند . در اين حالت سلول داراي حداقل تحريك پذيري است . جزء ديگر گروه keir كانال پتاسيم حساس ATP است . اين نوع كانال نمونه اي است كه چگونه متابوليسم و تحريك پذيري مي توانند با هم در ارتباط باشند . از نظر متابوليكي ، فیبرهای عضله تحليل رفته كاهش در غلظت ATP سيتوسوليك به علاوه در PH ناشي از افزايش زياد رسانايي پتاسيم در غشاء است كه به علت بازشدن كانالهاي پتاسيم حساس ATP اتفاق مي افتد اينگونه فرض مي شود كه در ارتباط با مصرف ناكافي پتاسيم از طريق na-k-Atpuse باعث كاهش ظرفيت غشاء سلولي براي توليد پتانسيل فعال همراه با كاهش نيرو مي شود.
توزيع و انتشار نامساوي كاتيونهاي يك ظرفيتي در سراسر غشاء پلاسما از طريق توليد و حفظ مي شود . اين پمپ محرك ATP الكتروزيك است ، در طي هر سيكل و چرخه تنها دو يون پتاسيم براي تبادل سه يون سديم خارج شده ، وارد مي شوند . پمپ سديم – پتاسيم شامل دو زير واحد است :
زير واحد كاتاليزور و زيرواحد گليكوپروتئين ايزوفرمهاي متعددي از پمپ هاي وجود دارد در حال حاضر چهار ايزوفرم و سه ايزوفرم گليكوپروتئين B شناسايي شده است در بافت هاي عضلاني وجود ايزوفرمهاي بستگي به نوع فيبرها دارد . در جوشهاي صحرايي ، ايزوفرمهاي غالب ماهيچه اي اكسيداتيو قرمز هستند در حاليكه در ماهيچه هاي
گليكوليتيك سفيد تنها ايزوفرمهاي وجود دارند در ماهيچه ها با نيروهاي تركيبي هم زيرواحدهاي بافت مي شوند.
در طي شرايط ساكن فعاليت پمپ سديم – پتانسيل تنها مساوي با 5% از حداكثر ظرفيت پمپ در فيبرهاي مجزاي عضله است . با شروع فعاليت انقباض ميزان پمپ سديم – پتاسيم افزايش مي يابد تا گراديان يوني حفظ شود . ظرفيت پمپ سديم – پتاسيم اصولا از طريق دو مكانيزم مختلف قابل تقويت و افزايش است از طريق تعديل و تنظيم تعداد پمپ هاي فعال سديم و پتاسيم
در غشاء پلاسمايي و يا از طريق تقويت فعاليت پمپ هاي يوني در حقيقت جابجايي پمپ سديم – پتاسيم از جدارهاي داخل سلولي به سمت غشاء پلاسمايي اخيرا تشريح شده است (Hundal و همكارانش از سال 1992 ، lavaie وهمكارانش 1996 ، jual و همكارانش در 2000 ) علاوه بر اين فعاليت پمپ سديم از طريق مكانيزمهاي متعددي نظير غلظت سديم داخل سلول ، سطح AMP چرخه اي از طريق ساير پروتئين كيفاز A وجود ATP ، PH ، پتانسيل غشاء و فشار استرس قابل كنترل و تنظيم است . و اين مورد كه آيا افت چگالي وتراكم در ميزان ATP مرتبط با تحريك پذيري پمپ سديم – پتاسيم نامشخص و مبهم است به اين علت است كه K,S (غلظت ATP كه پمپ سديم – پتاسيم را تحريك مي كند تا 50% ميزان حداكثر پمپ است ) در حدود است و محدوده معمولا حتي در طي تمرينات ورزشي كامل به آن ميزان نمي رسد .
ناقلهاي مرتبط
ناقلهاي مشترك مرتبط (KNCC) متعلق به خانواده ناقلهاي كلرويد داراي ؟؟؟؟؟؟؟كاتيون هستند (CCC) . قسمت ديگري از اين خانواده ناقل كلرويد پتاسيم است كه بعدا مورد بحث قرار مي گيرد دو ايزو فرم از NKCC با جرم ملكولي در حدود
130KDA,122 شناسايي شده اند . پروتئين شامل منطقه مرکزی هيدروفوبيك با 12 محدوده ميان سلولي مرتبط يك آمينو و يك ناحيه ترمينال كربوكسي در كنار هم قرار گرفته اند . در صورتيكه تاكنون ،ايزوفرم NKCC2 تنها در كليه يافت مي شود ، ايزوفرم NKCCL از طريق گراديان شيميائي تركيبي يونهاي منتقل شده بصورت يك محرك ناقل فعال ثانويه قابل تقسيم بندي است .
با اين وجود ، شواهدي وجودي دارد كه عملكرد آن بستگي به قابليت دسترسي ، ATP دارد گرچه وضعيت فعال سازي ATP با ATP از نوع متفاوت است با توجه به اين نظريه كه ATP بر فعاليت NKCC از طريق مكانيزم فسفريلدار كردن بروي فسفريل دار كردن پروتئين تأثيرگذار است NKCC يونها را بطور الکتریکی به آرامي از سلول با استوشيومتري غیر معمول منتقل مي كنند گرچه حداقل در آسكون هاي قلابي شكل شواهدي در مورد استوشيومتري غير معمول وجود دارد . لوپ ادرار آور نظر bumetanide (از بازدارنده هاي مهم NKCC هستند . علاوه بر اين افزايش در غلظت كلريد داخل سلولي و PH اسيدي درون سلولي مانع NKCC است . از لحاظ عملكردي Nka شامل فرايندها و مراحل مختلف ترشحي وجذب كننده مخاطي است به علاوه شواهدي در مورد نقش مهم Nkcc در تنظيم حجم سلولي وجود دارد.
ناقل
چهار ژن مختلف شناسايي شده اند كه براي چهار ايزوفرم مختلف از ناقل رمزگذاري شده اند . در ميان تنها ايزوفرم kcc1 بطور موجود در بافت هاي مختلف توزيع و منتشر شده است . شبيه ناقل ناقل يك محرك ثانويه ناقل يون هاي از طريق گراديان يونهاي جابجا شده است بيشترين ويژگيها و
مشخصه هاي ناقل در سلول هاي قرمز خوني به نام انتقال الكتريكي خنثي نيروي جاذبه يونها ، انرژي جنبشي و بازدارنده دارويي متمايز شده اند . افزايش در غلظت كلريد درون سلولي فعاليت ناقلهاي مرتبط را تحريك مي كند . علاوه بر اين انواع زيادي ار اكسيدانت ها مي توانند ناقلهاي را نظير H2O و دي اكسيد را تحرك كنند . مشابه با موقعيت NKCC ، فعال سازي ناقل كلريد پتاسيم از طريق PH,ATP تنظيم و تعديل مي شود از نظر عملكردي ناقل شامل تنظيم و كنترل حجم سلولي ، تعادل يوني درون سلولي است . تحقيقات اخير نيز نقش اين ناقل را در كنترل مقاومت شيرياني پيراموني را ارائه مي دهد.
تعادل و توازن پتاسيم درون سلولي :
توزيع و انتشار پتاسيم ميان جداره هاي داخل و خارج سلولي پتانسيل ساكن غشاء و تحريك پذيري سلولي را تعيين و مشخص مي كند . غلظت پتاسيم در مايع خارج سلولي وجود دارد .
ترشح پتاسيم از طريق كليه و با حداقل ميزان ترشح معده و روده اي ترشح پتاسيم از طريق عرق كه تنها در شرايط وزشهاي شديد جسماني نقش مهمي دارد.
تغيير ميزان پتاسيم ميان جداره داخلي و خارجي سلول . براي جزئيات بيشتر درباره تنظيم پتاسيم خارج سلولي خواننده به كسب فيزيولوژي و بررسيهاي اخير (1998 kamel,Halperin ,1999 Giebisch) مراجعه كنند.
تقريبا 8% از ميزان پتاسيم بدن در جداره داخل سلولي وجود دارد ، نقش تنظيم پتاسيم داخل سلولي در اين قسمت مورد تأكيد و اهميت قرار مي گيرد.
غلظت پتاسيم سلولي تابع يك تنظيم كوتاه مدت و طولاني است همانطور شكل 6-1 در حاليكه تاثيرات تمرينات سخت و شديد كه بعدا به آن پرداخته مي شود . اگر فاكتورهاي
فایل : 34 صفحه
فرمت : Word
- کاربر گرامی، در این وب سایت تا حد امکان سعی کرده ایم تمام مقالات را با نام پدیدآورندگان آن منتشر کنیم، لذا خواهشمندیم در صورتی که به هر دلیلی تمایلی به انتشار مقاله خود در ارتیکل فارسی را ندارید با ما در تماس باشید تا در اسرع وقت نسبت به پیگیری موضوع اقدام کنیم.


 454875 کاربر
454875 کاربر  85472455
85472455  4512145
مطلب
4512145
مطلب  3564215 دیدگاه
3564215 دیدگاه 

{kind=link}